The Lord Geekington Blog Posts
Will Brandt applied for and received a grant from the Web Archive Project enabling him to restore and archive this site for use by educators. It's a great example of how an interesting science based blog can credibly present material with attributions to all facts and claims. TNG/Earthling's Bob Sakayama managed the restoration along with Rev Sale who handled the development work. WAP provided funding for all initial work plus 5 years of hosting. Other team members from TNG provided research and technical support. The site has been referenced multiple times by both secondary and college level courses as an example of content presentation and for the facts provided by the content. Visitors are encouraged to copy and share all content.
For a number of years Cameron McCormick posted Cryptozoological Case Files for his followers on The Lord Geekington.
Content below are some of the 2011 Blog posts providing a small example of what was offered.

The Lord Geekington
Cameron McCormick
A college graduate - now what?
Saturday, December 17, 2011
Feresa: The Growling Wolf-Dolphin
 |
| Feresea... maybe. This species can be distinguished from Pseudorca by having a proportionally larger dorsal fin (2 base lengths away from the blowhole vs. 2.5) and by having a clearly demarcated cape; Peponocephala can be distinguished by having pointed flipper tips, a pointed head when viewed from above, and no white extending around the face (Baird 2010). I think this is the case in the above photo, but I'm not entirely certain. Photo by Gary L. Friedrichsen from WoRMS. |
In 1965 - a little over a decade after the external appearance of the animal became known - Feresa was held in captivity. Pryor (1991) remarked that one individual behaved "more like a wolf than a normal dolphin" would "growl and snap like as canid" and "not hesitate to attack people and other cetaceans". Since when are cetaceans capable of growling? This behavior has led some to presume that Feresa preys on mammals in the wild (Leatherwood et al. 1982) and aggression towards other dolphins has been observed whilst individuals were trapped in tuna seines (Reeves et al. 2002). Considering that both situations occurred in cramped and undoubtedly stressful environments, I think it is completely unfounded to conclude that Feresa is a pugnacious marine mammal-killing macropredator with the available evidence. Stomach contents have included squid and fish (Rodríguez-López and Mignucci- Giannoni 1999; Zerbini & Santos 1997)
 |
| Feresa skeleton. From Wikipedia Commons. |
References:
Baird, R. W. (2010). Pygmy Killer Whales (Feresa attenuata) or False Killer Whales (Pseudorca crassidens) Identification of a Group of Small Cetaceans Seen off Ecuador in 2003. Aquatic Mammals 36(3), 326-327.
Gray, J. E. (1871). Supplement to the Catalogue of seals and whales in the British Museum. Available.
Leatherwood, S., Reeves, R. R., Perrin, W. F., & Evans, W. (1982). Whales, dolphins, and porpoises of the eastern north pacific and adjacent arctic waters (NOAA Technical Report NMFA Circular 444). Washington, DC: U.S. Department of Commerce, National Oceanic and Atmospheric Administration, National Marine Fisheries Service. Partially
McSweeney, D. J., Baird, R. W., Mahaffy, S. D., Webster, D. L., and Schorr, G. S. (2009). Site ï¬delity and association patterns of a rare species: Pygmy killer whales (Feresa attenuata) in the main Hawaiian Island. Marine Mammal Science 25(3), 557-572.
Pryor, K. (1991). Mortal remains: Studying dead animals. In: Pryor, K. & Norris, K. S. (eds.) Dolphin Society: Discoveries and Puzzles. University of California Press: Berkeley.
Reeves, R. R., Stewart, B. S., Clapham, P. J., & Powell, J. A. (2002). National Audubon Society Guide to Marine Mammals of the World. Alfred A. Knopf: New York.
Rodríguez-López, M. A. & Mignucci-Giannoni, A. A. (1999). A stranded pygmy killer whale (Feresa attenuata) in Puerto Rico. Aquatic Mammals 25(2), 119-121. Available.
Rossi-Santos, M., Baracho, C., Neto, E. S., & Marcovaldi, E. (2006). First sightings of the pygmy killer whale, Feresa attenuata, for the Brazilian coast. JMBA2 - Biodiversity Records.
Slater, G. J., Price, S. A., Santini, F., and Alfaro, M. E. (2010). Diversity versus disparity and the radiation of modern cetaceans. Proceedings of the Royal Society B 277(1697), 3097-3104.
Vilstrup, J. T., Ho, S. Y. W., Foote, A. D., Morin, P. A., Kreb, D., Krützen, M., Parra, G. J., Robertson, K. M., de Stephanis, R., Verborgh, P., Willerslev, E., Orlando, L., & Gilbert, M. T. P. (2011). Mitogenomic phylogenetic analyses of the Delphinidae with an emphasis on the Globicephalinae. BMC Evolutionary Biology 11(65).
Zerbini, A. N. & Santos, M. C. O. (1997). First record of the pygmy killer whale Feresa attenuata (Gray, 1874) for the Brazilian coast. Aquatic Mammals 23(2), 105-109.
Thursday, December 15, 2011
Eocetus, "Eocetus", and Friends
|
|
| Lumbar vertebrae in right lateral view. From left to right: "Eocetus" wardii (from Uhen 1999), Basiloterus hussaini, and Basilosaurus drazindai - note that the latter-most may be an anterior caudal (from Gingerich et al. 1997). For comparison: Basilosaurus isis vertebrae. |

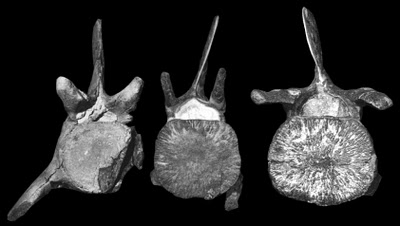 |
| Lumbar vertebrae in anterior view. Ditto order. |
As for awkward middle-child Basiloterus, it appears to have a centrum which is slightly more elongated than that of "Eocetus", however the neural arch and maybe the neural spine appear to be narrower. The metapophyses are upwardly-angled (Gingerich et al. 1997), less broad, less anterior-projecting, but still appear to extend past the centrum. The posterolateral processes are absent (Gingerich et al. 1997). The base of the transverse process is 9.3 cm long relative to a 19.5-20 cm centra (Gingerich et al. 1997), proportionally similar to Basilosaurus drazindai. The placement of Basiloterus is thus not clear, and perhaps it was a basilosaurid or an even more derived protocetid.
 |
| Maiacetus inuus, a basal "protocetid" (Uhen 2011). From Wikipedia Commons. |
 |
| Dorudon atrox. From Wikipedia Commons. |
The scare quotes around "Eocetus" hint at a taxonomic misadventure. "E." wardii was assigned to its genus by Uhen (1999) based on comparisons of its skull and vertebrae to Eocetus schweinfurthi; the problem is, the holotype of E. schweinfurthi is an isolated skull and it is not possible to determine whether the vertebrae referred to it actually represent the species (Geisler et al. 2005). There is overlapping skull material (Uhen 1999), but Geisler et al. (2005) apparently regarded it as too incomplete to warrant unambiguous placement in the genus. Somehow, "Eocetus" and Eocetus formed a clade in phylogenetic analyses (Geisler et al. 2005, Uhen et al. 2011), making it probable that future discoveries will confirm their close relationship.
"Eocetus" wardii is clearly related to unnamed Pisco Formation material which exhibits the same distinctive traits (moderate centrum elongation, elongated neural arches and spines and transverse processes, strange pock-marked texture) with the only difference being that the unnamed material is 35% smaller (Uhen et al. 2011). The Egyptian vertebrae dubiously assigned to Eocetus schweinfurthi (figured in Uhen 1999) also seem quite similar (including the pock-marks), and if it is also a member of this clade, it would indicate a sizable trans-oceanic range. This in turn could be taken as evidence of the whales being largely pelagic... of course this is quite speculative.
There of course remains much to be known about these cetaceans, and perhaps future discoveries will be enlightening as to how similar they were to the pelagic cetaceans, as well as the origins of Pelagiceti. I really hope it turns out that a Basilosaurus-sized animal could walk on land.
References:
Geisler, J. H., Sanders, A. E., and Luo, Z-X. (2005). A New Protocetid Whale (Cetacea: Archaeoceti) from the Late Middle Eocene of South Carolina. American Museum Novitates 3480, 1-65..
Gingerich, P. D., Arif, M., Bhatti, M. A., Anwar, M., & Sanders, W. J. (1997). Basilosaurus drazindai and Basiloterus hussaini, new Archaeoceti (Mammalia, Cetacea) from the middle Eocene Drazinda Formation, with a revised interpretation of ages of whale-bearing strata in the Kirthar Group of the Sulaiman Range, Punjab (Pakistan). Contributions from the Museum of Paleontology, University of Michigan 30 (2), 55-81. .
Uhen, M. D., Pyenson, N. D., Devries, T. J., Urbina, M., and Renne, P. R. (2011). New middle Eocene whales from the Pisco Basin of Peru. Journal of Paleontology 85(5), 955-969. doi: http://dx.doi.org/10.1666/10-162.1
Uhen, M. D. (2010). The Origin(s) of Whales. Annual Review of Earth and Planetary Sciences 38, 189–221.
Uhen, M. D. (2008). Basilosaurids. In: Perrin, W. F., Würsig, B., and Thewissen, J. G. M. (eds.) Encyclopedia of Marine Mammals, Second Edition. Elsevier: Burlington, Massachusetts.
Uhen, M. D. (1999). New Species of Protocetid Archaeocete Whale, Eocetus wardii (Mammalia: Cetacea) from the Middle Eocene of North Carolina. Journal of Paleontology 73(3), 512-528.
Weems, R. E., Edwards, L. E., Osborne, J. E., and Alford, A. A. (2011). An occurrence of the protocetid whale "Eocetus" wardii in the Middle Eocene formation of Virginia. Journal of Paleontology 85(2), 271-278.
Wednesday, November 30, 2011
Billfish Bills - What Are They Good For?
In the prior article, I discussed long-beaked "dolphins" (Eurhinodelphidae) and noted that I couldn't find hypotheses on the function of their uneven jaws in the literature... aside from a weird proposal involving Skimmers. The Theatrical Tanystropheus mentioned a couple ("digging for small, sand-dwelling organisms or as a bat with which to stun fish") which are plausible, but I don't know where they are from or what lines of reasoning are behind them. There are extant species with a superficially similar condition - billfishes - and it could be relevant to review what they do with their bills.
 |
|
Atlantic White Marlin (Tetrapturus albidus) from Wikipedia Commons. |
Swordfish (Xiphiidae) and Marlins/Sailfishes/Spearfishes (Istiophoridae) are living sister taxa1 in the clade Xiphioidea; while traditionally included in Scombroidei, billfishes are presently regarded as phylogenetically distinct (Orrell et al. 2006) and possibly close relatives of jacks and... flatfishes (Little et al. 2010). Fish phylogenetics is scary business, and I suspect billfish relations will undergo further revisions as the monstrosity known as "Perciformes" is reasoned into pieces. Anyways, while xiphiids2 and istiophorids look superficially similar, they actually have rather distinctive morphology. Swordfish have a bill which is flat in cross-section, toothless, blunt-tipped, and with central chambers (compared to rounded, denticulated, pointed, and chamber-less for istiophorids), a weak mandible much shorter than the rostrum, no scales, and no pelvic fins (Collette et al. 2006; Fierstine 2006; Fierstine and Voight 1996 citing Nakamura 1983). Strangely, most extinct billfishes have jaws of equal length, and if the proposed (Istiophoridae + Hemingwayidae) and (Xiphias + Xiphiorhynchus) clades (Fierstine 2006) are correct (see note 1), this would mean the unequal jaws of extant billfishes evolved twice.
1 A detailed cladistic analysis with the fossil members of the group has yet to be undertaken (Fierstine 2006).
2 As for what the deal with them and ziphiids is, I have no idea.
 |
| Swordfish (Xiphias gladius) from Wikipedia Commons. |
Fierstine (2006) hypothesized that unequal jaw length in billfishes may have evolved to avoid suffocation when impaling large objects (predator or prey) and to avoid damage to the mandible. I don't buy the mandibular reasoning since extant billfishes get by just fine with them naturally foreshortened. The available evidence suggests impaling is a rather rare event and thus unlikely to be the main factor in the evolution of the characteristic billfish bill. An alternate hypothesis could be that the mandible was shortened so the rostrum could be "weaponized" (sword-like flattening in xiphiids and denticles in istiophorids3) to slash at prey. However, the population which apparently doesn't use bills to feed and healthy individuals with damaged rostra are problematic for both of these hypotheses. Perhaps future studies will show that the bill is generally important for feeding in the group and that the counterexamples are just freaks, but either way, it seems premature to make any conclusions about why billfish have their striking morphology.
The ichthyosaur Eurhinosaurus has teeth on the upper jaw which could be a similar instance of "weaponization".
I really have no idea how eurhinodelphids fit into this framework since Fierstine's hypothetical suffocation would not be an issue (if they could impale at all) and the rostrum does not seem particularly dangerous (no teeth, denticles, or flattening). I wonder if this morphology evolved for different reasons, or if it evolved for reasons that have yet to be hypothesized.
References:
Collette, B. B., McDowell, J. R., and Graves, J. E. (2006). Phylogeny of Recent Billfishes. Bulletin of Marine Science 79(3), 455-468.
Fierstine, H. L. (2006). Fossil history of Billfishes (Xiphioidea). Bulletin of Marine Science 79(3), 433-453.
Fierstine, H. L. (1997). An Atlantic Blue Marlin (Makaira nigricans), impaled by two species of billfishes (Teleostei: Istiophoridae). Bulletin of Marine Science 61(2), 495-499.
Fierstine, H. L., and Voight, N. L. (1996). Use of Rostral Characters for Identifying Adult Billfishes (Teleostei: Perciformes: Istiophoridae and Xiphiidae). Copeia 1996(1), 148-161.
Frazier, J. G., Fierstine, H. L., Beavers, S. C., Achaval, F., Suganuma, H., Pitman, R. L., Yamaguchi, Y., and Prigioni, C. M. (1994). Impalement of marine turtles (Reptilia, Chelonia: Cheloniidae and Dermochelyidae) by billfishes (Osteichthyes, Perciformes: Istiophoridae and Xiphiidae). Fisheries Science 39(1), 85-96.
Little, A. G., Lougheed, S. C., and Moyes, C. D. (2010). Evolutionary affinity of billfishes (Xiphiidae and Istiophoridae) and flatfishes (Plueronectiformes): Independent and trans-subordinal origins of endothermy in teleost fishes. Molecular Phylogenetics and Evolution 56(3), 897-904.
Nakamura, I. (1983). Systematics of billfishes (Xiphiidae and Istiophoridae). Publications of the Seto Marine Biological Laboratory 28, 255-396.
Orrell, T. M., Collette B. B., and Johnson, G. J. (2006). Molecular data supports separate scombroid and xiphioid clades. Bulletin of Marine Science 79(3), 505-519.
Shimose, T., Yokawa, K., Saito, H., and Tachihara, K. (2007). Evidence for use of the bill by blue marlin, Makaira nigricans, during feeding. Ichthyological Research 54(4), 420-422.
Vaske, T., Travassos, P. E., Pinheiro, P. B., Hazin, F. H. V., Tolotti, M. T., and Barbosa, T. M. (2011). Diet of the Blue Marlin (Makaira nigricans, Lacepède 1802) (Perciformes: Istiophoridae) of the southwestern equatorial Atlantic Ocean. Brazilian Journal of Aquatic Science and Technology 15(1), 65-70.
Posted by Cameron McCormick at Wednesday, November 30, 2011
Tuesday, November 29, 2011
Picture of the Indiscriminate Interval #000008 - Eurhinodelphis longirostris
 |
| Eurhinodelphis longirostris at the American Museum of Natural History. |
The most striking trait of Eurhinodelphidae is a toothless extension of the rostrum beyond the mandible (Lambert 2005), superficially similar to the bills of Billfish and Swordfish. Oddly, this morphology was speculative for a period of time (Kellogg 1925) although it has apparently been confirmed in several species as of Lambert (2005). Unfortunately, information on eurhinodelphids is scant and/or difficult to access and, among numerous other basics of their biology, I really don't know what the function of the extended rostrum would be. The only suggestion I could find is from one professor Abel who speculated that the cetaceans "swam on the surface of the sea, where they captured food - probably fishes - in much the same manner as does the skimmer (Rhynchops) [sic] among birds" (Anonymous 1909). Somehow, I find this even less plausible than azhdarchids-as-skimmers. On a curious note, there is a cetacean with the reverse of eurhinodelphid morphology (mandible extending past rostrum) unofficially known as the... skimmer porpoise
.
Phylogenetically, eurhinodelphids have bounced around from being considered stem-ziphiids, the sister group to Delphinida, and the sister group to Squalodontidae + Squalodelphidae (Geisler et al. 2011 - citing various); within Geisler et al. (2011), they were placed outside crown-Odontoceti1 in an unconstrained analysis and as the sister group of platanistoids in a constrained analysis. The authors regarded the latter position as more probable and placed eurhinodelphids within the new group Synrhina, which includes most odontocetes except for Sperm Whales and assorted extinct taxa. Whatever their placement, eurhinodelphids are certainly close relatives of living toothed whales, despite that whole extinct thing.

seems to have an unusually long neck for a cetacean. The cervical vertebrae are not fused (Kellogg 1925), however this is a surprisingly common trait shared with river dolphins, monodontids, rorquals, and gray whales (Tinker 1988). The neck of E. longirostris appears to be proportionally longer than those of the baleen whales and Narwhal and is probably comparable to those of the Beluga and Dorudon. River dolphin skeletons are hard to find, but it seems likely they have similarly proportioned necks. It seems that Eurhinodelphis wasn't a total freak, well, except for the snout.
The Theatrical Tanystropheus covered Eurhinodelphis as well, and it doesn't even overlap that much!
References:
Anonymous. (1909). Notes. Nature 2088 (82), 16.
Geisler, J. H., McGowen, M. R., Yang, G., Gatesy, J. (2011). A supermatrix analysis of genomic, morphological, and paleontological data from crown Cetacea. BMC Evolutionary Biology 11 (112).
Kellogg, R. (1925). On the occurrence of fossil porpoises of the genus Eurhinodelphis in North America. Proceedings of the U. S. National Museum 66(26), 1-40.
Lambert, O. (2005). Les dauphins longirostres et les baleines à bec du Néogène de la région d’Anvers: systématique, phylogénie, paléo-écologie et paléo-biogéographie. Doctoral Thesis
Tinker, S. W. (1988). Whales of the World. E. J. Brill Publishing Company: New York.
Friday, November 25, 2011
Picture of the Indiscriminate Interval #000007 - Narwhal
 |
| Monodon monoceros at the American Museum of Natural History. |
Narwhals are a bit strange even by cetacean standards. I'll let the title of this Tet Zoo article speak for itself: "A 3-m tooth that can bend 30 cm in any direction and is hypersentitive to salinity, temperature and pressure... and the sonic lance hypothesis".

They get weirder. Without the tooth (or sometimes, teeth) it is difficult to picture how this flat-skulled cetacean could be the same as a bulbous-headed Narwhal. As brought up in my Dorudon post, there are colossal amounts of soft tissue involved.

Another soft-tissue feature not hinted at by the skeleton are unusually shaped flukes... in males. Fontanella et al. (2010) suggest that the concave leading edge and lack of sweepback of the flukes increases lift and thrust to compensate for the drag caused by the tusk in males. The implications of the occasional tusked female narwhal were not discussed by the authors.


References:
Fontanella, J. E., Fish, F. E., Rybczynski, N., Nweeia, M. T., & Ketten, D. R. (2010). Three-dimensional geometry of the narwhal (Monodon monoceros) flukes in relation to hydrodynamics. Marine Mammal Science 27(4), 889-898.
Garde, E., Heide-Jørgensen, M. P., Hansen, S. H., Nachman, G., and Forchhammer, M. C. (2007). Age-specific growth and remarkable longevity in Narwhals (Monodon monoceros) from West Greenland as estimated by Aspartic Acid Racemization. Journal of Mammalogy 88(1), 49-58.
George, J. C., Follmann, E., Zeh, J., Sousa, M., Tarpley, R., Suydam, R. Horstmann-Dehn, L. (2011). A new way to estimate the age of bowhead whales (Balaena mysticetus) using ovarian corpora counts. Canadian Journal of Zoology 89(9), 840-852.doi: 10.1139/z11-057
George, J. C., and Bocktoce, J. R. (2008). Two historical weapon fragments as an aid to estimating the longevity and movements of bowhead whales. Polar Biology 31(6), 751-754.
George, J. C., Bada, J., Zeh, J., Scott, L., Brown, S. E., O'Hara, T., & Suydam, R. (1999). Age and growth estimates of bowhead whales (Balaena mysticetus) via aspartic acid racemization. Canadian Journal of Zoology 77, 571-578.
Posted by Cameron McCormick at Friday, November 25, 2011
Friday, November 18, 2011
The Giant Turtle Therizinosaurus
 |
| Therizinosaurus, you look... unwell. Reconstruction by K. K. Fierova, from Maleyev (1954). |
I am quite fond of old, weird reconstructions, and the initial classification of Therizinosaurus cheloniformis as a "turtle-like reptile"1 resulted in the magnificent specimen above. So how could the veritable Jabberwocky we're all familiar with be misinterpreted to such a colossal degree?
1 This odd phrasing is mirrored in the scientific name ("saurus" = lizard, "cheloniformis" = turtle-like). Malayev (1954) linked Therizinosaurus with members of Protostegidae and thus (probably) didn't intend to suggest another clade of reptiles which converged on turtles. Bizarrely, Rozhdestvensky (1974) claimed Malayev/Maleev classified Therizinosaurus as a "turtle-like pangolin"! Rozhdestvensky (1977) does not reiterate that statement, and further notes that another worker (Sukhanov) classified Therizinosaurus as a turtle; I unfortunately cannot find that source ("The subclass Testudinata" in Osnovy Paleontologii).
 |
|
Therizinosaurus in its non-turtle form. From Wikipedia Commons . |
Malayev (1954) described Therizinosaurus from scrappy remains: a metacarpal fragment, 3 manual unguals, and rib fragments (Zanno 2010). One of the ribs was an estimated 1.5 meters long when complete and was used to calculate a maximum body width of 3.25 meters (10'8") and body length of 4.5 m (14'9") (Malayev 1954); this is of course quite a bit larger than even the largest known Stupendemys geographicus.The rib was noted to lack costal elements, which is curious since turtle skeletons generally look like this:
 |
| Common Snapping Turtle (Chelydra serpentina) skeleton. Note the plastron is missing.
From Wikipedia Commons. |
Surprisingly, this is not necessarily a critical flaw, as (all?) turtles have distinct ribs during development before the carapace is fully formed (Wyneken 2001, fig. 90; Sánchez-Villagra 2009, figs. 3, 4). Malayev (1954) did not mention this nor the obvious possibility of a multi-ton hatchling. Instead, the "form of the ribs" was compared to Archelon and Protostega:
 |
| Archelon skeleton.
From Wikipedia Commons. |
The similarity is very general and Malayev (1954) does not list any specific shared characteristics. Due to the lack of costal elements, Malayev (1954) speculated that Therizinosaurus was in a distinct clade and in life had "barely developed or almost completely absent bony armor". It is incredibly strange that the Leatherback Seaturtle (Dermochelys coriacea) was not mentioned, as it entirely lacks costal elements and instead has thousands of dermal ossicles (Cebra-Thomas et al. 2005). The skeleton (sans ossicles) looks like an attempt by turtles to become "normal" tetrapods again.. until you notice the pectoral girdle within the ribcage:
 |
|
From Wikipedia Commons. |
The rib material used to describe Therizinosaurus cheloniformis is apparently not from a therizinosaur at all, but a sauropodomorph (Zanno 2010 citing Rozhdestvensky 1970). Isn't it a major problem that the holotype is a chimera? Whatever the case, Therizinosaurus cheloniformis has been re-described a few more times and other rib material has been referred to the species (Zanno 2010). However, all of the diagnostic traits (and most of the material) are from the forelimbs (Zanno 2010
Malayev (1954) interpreted the metacarpal and phalanges to be "powerful swimming organs" and suggested the huge claws were used for "cutting aquatic vegetation or for another functions, constrained by movement and acquiring food". The longest phalanyx was 60-65 cm long, not including the keratin covering (Malayev 1954), which suggests that the claws were ridiculously huge in life, even for a turtle-like reptile with a 4.5 meter body. I have observed turtles using their claws to climb and tear apart food (maybe what Malayev had in mind...), but clearly claws this disproportionate were doing something special. Something like this:
I like to think that Therizinosaurus, despite not being turtle-shaped anymore, waved its giant claws seductively in the faces of prospective mates.
References:
Cebra-Thomas, J., Tan, F., Sistla, S., Estes, E., Bender, G., Kim, C., Riccio, P., and Gilbert S. F. (2005). How the Turtle Forms its Shell: A Paracrine Hypothesis of Carapace Formation. Journal of Experimental Zoology 304B, 558-569.
Maleyev, E. A. (1954). A new turtle-like reptile from Mongolia. Priroda 3, 106-108.
Rozhdestvensky, A. K. (1977). The study of Dinosaurs in Asia. Journal of the Palaeontological Society of India 20, 102-119.
Rozhdestvensky, A. K. (1974). History of the dinosaur fauna of Asia and other continents and questions concerning paleogeography. Transactions of the Joint Soviet–Mongolia Paleontological Expedition 1, 107–131.
Rozhdestvensky, A. K. (1970). On the gigantic claws of mysterious Mesozoic reptiles. Palaeontological Journal 1, 131-141.
Sánchez-Villagra, M. R., Müller, H., Sheil, C. A., Scheyer, T. M., Nagashima, H., and Kuratani, S. (2009). Skeletal Development in the Chinese Soft-Shelled Turtle Pelodiscus sinensis (Testudines: Trionychidae). Journal of Morphology 270, 1381-1399.
Wyneken, J. (2001). The Anatomy of Sea Turtles. U.S. Dept Commerce NOAA Tech Mem NMFS SEFSC-470.
Zanno, L. E. (2010). A taxonomic and phylogenetic re-evaluation of Therizinosauria (Dinosauria: Maniraptora). Journal of Systematic Palaeontology 8(4), 503-543.
Posted by Cameron McCormick at Friday, November 18, 2011
Tuesday, November 8, 2011
A Giant Snapper At Last!
Of course, I've documented a big snapping turtle before, but my subsequent failures were astounding. I saw the turtles on multiple occasions this year (alluded to here), sometimes up close (touching, in fact) and once in triplicate. On all of these occasions I didn't bring my camera because I was commuting via bike, or the turtles fled before I could get their photographs. After a couple dozen failures, I gave up. Impulsively I decided on 8 November 2011 to take a trip searching for any reptiles or amphibians still active in the abnormally warm weather (about 70° F, 21° C) and saw this (plus a frog):


Getting closer, I confirmed my suspicions that, yes, this turtle is really really big.

The closest thing I could get to a measurement.

I know I'm never going to get an accurate length or weight measurement from this turtle... not without one or both of us getting hurt. Even if this specimen was a record (and there's no guarantee), it would not be worth risking the life of an old reptile to revise the SCLmax of 49.4 cm for Chelydra serpentina. From now on I'm leaving these turtles alone, my curiosity is satiated, and bothering them further will have no benefits. I'll have to live with the wonder that despite inhabiting a polluted body of water and having human hunt them and compete for their resources, things like this still exist.